碳水化合物酶在猪日粮中的使用,是基于粘性谷物(如小麦和大麦等)在日粮中的使用,与家禽日粮中使用酶制剂是同期的,但是直到最近,在猪日粮中的应用研究还远不能使人们信服。造成这种情况有很多原因,当然并非所有原因与投入产出(这是最终是否采纳一种新的饲料技术的关键)有关。在很多国家养猪和养禽业的基本结构(如高度集约型与半粗放型相比)的不同,是养猪业迅速采纳新的饲养技术的限制性因素。这种情况(吸收新技术的潜能)在世界上有些国家的猪市场上正在发生快速的变化,毋庸置疑这将促进养猪也向着更好的方向发展。
因为植酸酶以及直接作用于植物蛋白饼粕的酶制剂,在本书的提他章节一有详细论述,所以,本章将要讨论,碳水化合物酶在谷物基础日粮中的作用和使用效果。在着手写本章内容时,饲料酶制剂在猪日粮中的使用,还落后于家禽,但是这种情况会迅速变化,尤其是在过去的5年,已经后更多的更有效的,在猪日粮中使用的酶制剂上市,和更多的应用研究。因为有这种迅速的变化,所以本将要综述过去10年里在本领域发表的文献。
对酶的反应:猪与禽的比较 有很多学者在很多偶然的场合谈到,猪和家禽对日粮中添加外源性碳水化合物酶的反应不同的原因可能其生理解剖结构的不同引起的,因此这里将总结这些原因。在过去的10年里,读者可能会看到很多著名的研究报告,系统地报道了这些原因以及酶制剂在猪日粮中使用方面的文献。这些文献中有些是把猪和家禽进行比较或对比(如Dierich,1989;Chesson,1993;Dierick和Decuypere,1994,1996;Graham和Balnave,1995;Berdford和Schulze,1998;Danicke等,1999)。
与家禽相比,影响猪对酶制剂的潜在反应的主要因素有:胃肠道解剖结构、消化能力以及食糜的特征等。
因为外源性酶制剂的最佳pH不同,肉仔鸡的嗉囊存在使得一些酶在进入pH值低的肌胃之前,首先在相对高的pH值环境(约pH=6)中发挥活性。而对于猪来讲,在胃的pH值更低的环境中(pH<3),酶制剂会迅速与其底物接触,尽管胃pH值受到猪的年龄、饲料原料的缓冲程度以及胃中微生物发酵产生有机酸(如:乳酸)的程度的影响,但还是很低。因为猪的胃肠蠕动相对缓慢(Moran,1982),而导致胃中不同区域的pH值以及内源酶活性不同。所有的这些因素均会因pH值、温度特性以及粘附反应常数(Km),而潜在地影响到外源性酶的活性。
家禽的固态和液态食糜的平均保留时间(MRTs)比猪少(Moran,1982)。肉仔鸡胃肠道上部(嗉囊、肌胃和小肠)可溶性标记物的MRT分别为2.8、0.3和1.0小时(约需要4个小时到大小肠后部,Dierick和Decuypere,1994)。Dierick和Decuypere(1994)证实食糜通过猪为和小肠的MRT为4-5个小时,而Mahan(1982)研究表明此时间为7.5小时。两种动物的日粮可溶性物质含量均会因日粮组成、可溶/不可溶纤维水平、饲养方式、动物年龄等而变化,但是因为肉仔鸡对营养有效消化和吸收的时间比猪短,因此潜在地受特定抗营养因子(如日粮粘度、高分子量的可溶性纤维)的影响更明显。所以两种动物对外源性酶制剂的需要及相对重要性不同,尽管其消化道的长度以及MRT不同,但是两种动物均缺乏消化这些抗营养因子的适当酶。因此,两种动物均受到同样的消化紊乱的影响,不仅是营养物质吸收减少,而且小肠细菌数量会增加,从而导致宿主与细菌之间对营养物质的竞争。
与肉仔鸡相比,细菌在猪肠道中的繁殖数量较大,这也是通常认为的影响猪对外源性酶制剂的反应的原因。当然食糜在猪大肠中存留时间长,盲肠和直肠的体积较大,是细菌对残留纤维发酵的最佳场地,发酵产生的挥发性脂肪酸可以满足猪的维持需要。但是正如前文所提到的,小肠中细菌的大量繁殖并非好事。一些研究报道在日粮不添加外源性酶制剂时,猪小肠中纤维有显著的断裂。这些研究中有些使用了“T”漏管在猪回肠末端插入,也有使用十二指肠部分插入的(Graham等,1986,1988)。非常规菌在漏管区的定植,会最终影响试验结果。
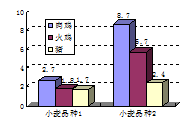
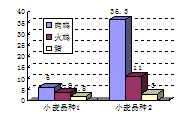
众所周知,粘性谷物的可溶性、大分子量、非淀粉多糖(NSPs)能干扰鸡对营养物质的消化和吸收,导致微生物在肠中的繁殖(Apajahti和Bedford,1998;Bedford和Schulze,1998)。猪肠道食糜的天然高的水分含量(比饲喂同样日粮的家禽高10%),会在一定程度上冲淡因食糜粘度问题而带来的副作用(图8.1;Danicke等,1999)。但这并不是说,在猪粘性谷物日粮中使用碳水化合物酶没有作用,而是说相对于家禽来说,碳水化合物酶对猪的重要性差一些。很多试验研究表明,添加碳水化合物酶可以降低饲喂粘性谷物日粮的猪的肠道食糜的粘度(图8.2),但是起始的粘度水平均显著低于家禽的试验结果。但本质上可溶性纤维是在疾病和饲喂特定日粮条件下,猪肠道中有害细菌变化的关键因素(本章后面将要讨论到),因此不能低估粘度对猪生产性能的影响。所以我们认识到,这种影响(通过对肠道微生物群落的间接影响)比直接影响营养物质的消化更温和。
图8.1(1)空肠的情况 图8.1(2)回肠的情况 途中纵坐标为小肠食糜粘度。小麦品种对肉仔鸡、火鸡和猪小肠粘度(mPa s)的影响(Danicke等,1999)。肉仔鸡(3周龄)日粮小麦添加量73%;火鸡(4周龄)日粮小麦添加量73%;猪(8周龄)日粮中小麦添加量86%*。*:小肠是指猪的下2/3小肠。小麦品种1=“Ibis”,可溶性阿拉伯木聚糖含量为11 g/kg干物质;小麦品种2=“Alidos”,可溶性阿拉伯木聚糖含量为17 g/kg干物质。
图8.2酶对仔猪小肠粘度的影响 参考文献1:Inborr(1994),p<0.05;参考文献2:Sudendey和Kamphues(1995),小麦/仔猪;大麦/生长猪,p<0.05;参考文献3:Dusel等(1997),p<0.05;参考文献4:Partridge等(1998a),p<0.01。
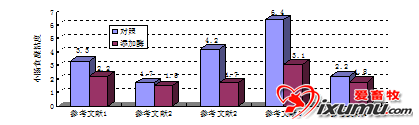
研究表明,向饲喂小麦或大麦基础日粮的仔猪活生长猪日粮中添加酶制剂后,猪胃部食糜干物质流速增加(Sudendey和Kamphues,1995;图8.3)。在这些试验中,饲喂酶制剂的动物的胃中的食糜粘度会迅速降低。Ellis等(1996)发现猪胃中食糜粘度的增加,会因为大的食糜颗粒不能沉到胃的底部而导致筛选机制失败,这种特征性现象可能是猪上唯一的。在粘性环境中,这些大颗粒仍悬浮着,在被胃分泌物消化或者食糜颗粒变少之前,更易逃出幽门,从而影响接下来的消化。进而在胃肠道中,粘性物质会影响胃肠的蠕动,构建层流区,比影响流动更厉害,也能通过影响胃抑多肽而影响胰腺的分泌。粘性非淀粉多糖业能物理性的包被淀粉颗粒,从而降低淀粉的消化率(Ellis等,1996)。
图8.3小麦或大麦基础日粮中添加酶制剂,对动物胃中干物质流速的影响(Sudendey和Kamphues,1995)。以每公斤体重每小时离开胃的干物质克数计流速。添加的酶制剂由淀粉酶、木聚糖酶和β-葡聚糖酶组成。小麦/仔猪组差异极显著(p<0.01);大麦/生长猪组差异显著(p<0.05)。 在使用酶制剂的成功猪试验中,猪日增重的增加很大部分是因为酶制剂的使用刺激采食量增加的结果(Haberer等,1998)。胃排空率和胃的膨胀是猪产生饱感的许多信号因素中的两个(Forbes,1995),这种情况似乎与实际情况不一致,如有人认为外源性酶制剂对增加采食量的正效应,可能是因为对各种因素对食糜流速的影响,以及对小肠中营养物质的消化和吸收的改善而致。正如Bedford和Schulze(1998)报道的那样,肠道激素的反馈回路,对碳水化合物酶的作用也有影响,他们认为盲肠中的发酵,会因木聚糖酶的添加增加了降解产物(可发酵寡糖)而增强,进而影响肠道糖原的浓度,从而通过降低胃泌素的浓度而影响胃的蠕动。这些可能性表明,外源性酶制剂能通过不同的机制有效影响胃肠道生理,但是这方面的研究较少。
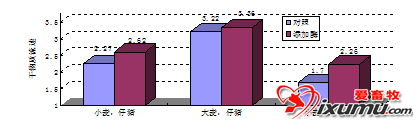
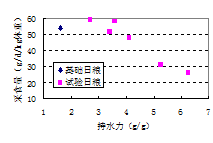
与肉仔鸡相比,猪肠道中食糜的通过率相对较低,可能更有利于外源性酶制剂的使用,随着食糜沿着小肠向下移动,可使不溶性细胞壁的破裂增加或变小,也能使纤维发生理化变化(如持水力降低)。Kyriazakis和Emmans(1995)发现对仔猪(12-25 kg)来说,饲料持水力是影响采食量的主要因素(图8.4)。因为体外试验表明碳水化合物酶可以降低饲料的持水力(图8.5),猪日粮中添加外源性酶制剂的一些益处,可能与肠道中残留的可溶性和不可溶性纤维打破了水与营养物质的嵌和有关。可溶性粘性、非淀粉多糖消化分解的重要性,可能使饲喂小麦和大麦的肉仔鸡生产性能改善的主要作用模式(Bedford和Schulze,1998),在猪上这种重要性意义不大。猪日粮中添加外源性碳水化合物酶制剂的作用机制可能很多,不仅局限于外源性酶制剂对肠道中可溶性纤维(尤其是不可溶性纤维)。这关系到产品设计,因为不同酶制剂对不同纤维结构的亲和性和反应速率不同,因此为猪设计最佳效率的酶制剂应该包括不同水平和/或不同来源的酶,相对来说为家禽设计酶制剂更简单一些。以木聚糖酶为例,甚至来自同一种微生物的木聚糖酶,对不同木聚糖底物的催化活性变异也很大(Bedford和Schulze,1998),木聚糖酶的活性浓度也是细胞壁水解程度的重要的决定性因素(Tervila-Wilo等,1996)。
图8.4 饲料持水力对猪(12-25 kg活体重)采食量的影响(Kyriazakis和Emmans,1995)。
为非粘性谷物(如玉米、高粱等,也是世界上使用最普遍的谷物)而设计的碳水化合物酶,是否能商品化,这种情况在当今显得尤其重要。在这些谷物中,与不可溶性纤维相关的因素,如细胞壁内对营养物质的包被、以及淀粉的结构和组成,与不可溶性纤维的相关性更大,需要一系列新型酶制剂来降解。
接下来本章将要系统地讨论,世界上在养猪业中使用的主要谷物及谷物副产品的来源,考查其饲养价值及变异程度,以及造成变异的原因。而后综述近些年来,利用酶制剂来提高其潜在营养价值的研究。
大麦作为猪日粮的谷物来源 Barneveld(1999)综述了传统(带壳)大麦对猪的饲养价值,从16个实验室的研究估计其消化能(DE)在11.7-16.0 MJ/kg干物质(DM)。能量消化率,尤其是总能含量(变异范围在±2 MJ/kg DM)对DE(消化能)值的影响最大。最近Fairbairn等(1999)研究了生长于3个地方的5个品种共20个大麦样品的DE含量,研究表明其DE含量在11.2-13.1 MJ/kg(90%DM)之间,与品种内的变异(8.4%,1.0 MJ/kg)相比,品种间的变异(5.8%,0.7 MJ/kg)较小。这再次强调了生长环境对组分分析、碳水化合物组成以及最终对猪和鸡的饲养价值的重要性(Aman等,1985;Aman和Newman,1986;Bach Knudsen等,1987)。
最近的10年人开始尝试用脱壳大麦作为单胃动物的饲料原料,脱壳大麦与传统的大麦(带壳)不同,壳在脱壳过程中被脱去,与去壳后的部分不再紧密相连。尽管脱壳使得大麦的不可溶纤维的含量降低、蛋白含量升高,但其对猪鸡的饲养价值却并未达到理论上应提高的程度(Baidoo等,1998b;Thacker,1999)。这可能与大麦的胚乳中大量存在的可溶性纤维部分(尤其是β-葡聚糖酶)有关。脱壳后使得这些可溶性纤维在脱壳大麦中的含量升高,如带壳大麦的β-葡聚糖酶含量为3.5%-4.5%,脱壳大麦的β-葡聚糖酶为4.5%-7%(Baidoo和Liu,1998)。饲喂生长/肥育猪微粉化的脱壳大麦后对生长性能的影响,是因为这种纤维的的可溶性进一步提高,尤其是在对这种谷物的热加工过程中,掩盖了对淀粉凝胶过程的促进作用(Thacker,1999)。Thacker(1999)的研究再次强调了,在生产性能的研究中测定猪总肠道消化率的局限性,营养物质“表观”消化率的改善,不仅因采食量降低,而且因影响微生物繁殖/对营养物质的竞争而抵消,这将在本章的后面部分详细论述。
带壳和脱壳大麦饲养价值的不同,也会受到除纤维外谷物中其他成分的影响,尤其是淀粉部分中直链淀粉与支链淀粉比例不同的影响。Pettersson和Lindberg(1997)以生长育肥猪为试验动物,研究了含不同比例直链淀粉与支链淀粉的带壳与脱壳大麦的回肠和粪消化率。在回肠和粪水平的研究均表明,带壳高直链淀粉含量的大麦的能量和淀粉消化率最低,脱壳支链淀粉含量高的大麦的能量和淀粉消化率最佳。与带壳大麦相比,脱壳大麦的营养物质消化率有更高的增加大肠消化的趋势。这大概暗示了微生物发酵率和发酵模式,但该研究没有涉及,也没有测定不同类型大麦对生产性能的影响,以确定消化率的不同实际在生长、采食量和饲料利用率上的表现。Miller等(1994)设计了消化试验,研究了纤维(ADF-酸性洗涤纤维)、β-葡聚糖酶含量以及直链淀粉与β-葡聚糖酶含量的比例之间的相互作用。研究表明大麦对猪和禽的能量含量随ADF含量增加而显著降低,但β-葡聚糖酶含量的增加对降低能量含量更明显。他们的研究还表明,β-葡聚糖酶对能量消化率的影响在禽比猪更明显,高支链淀粉(“蜡质”)含量的大麦淀粉抵消了一些因高ADF含量而对能量消化率的降低。这表明谷物不同部分之间的复杂的互作关系,会潜在的影响其饲养价值,对外源酶制剂会有所反映。
猪日粮中带壳和脱壳大麦对外源性酶制剂的反应 大麦的糊粉细胞壁中含有高水平的,由木聚糖围绕着一个复杂连接的β-葡聚糖(Autio等,1996)。因此,大麦基础日粮中同时添加木聚糖酶和β-葡聚糖酶时,动物才有最佳的生产性能。关于此点van Lunen和Schulze(1996a)以及Ramaswamy等(1996)的研究是很好的例证,他们的研究中带壳大麦基础日粮,只有在有较强木聚糖酶活性时,对动物才会有较好的饲喂效果。有些带壳大麦基础日粮中使用外源性酶制剂研究中(Graham等,1989;Thacker等,1992b),似乎仅强调了β-葡聚糖酶对降低可溶性β-葡聚糖(是肉仔鸡食糜粘度的主要成分)的作用。但在猪上类似作用的重要性较低,因为与肉仔鸡相比,猪食糜的干物质含量较低(Bedford和Schulze,1998),使得粘性纤维的浓度已经有效稀释,尤其是他们对营养物质消化和吸收的决定性影响。但是,粘性纤维仍通过对肠道微生物的影响而与此有关,因此而间接影响酶的活性,本章的下一部分将详细综述这方面的情况。
表8.1(见附表)总结了在过去的10年里,一些公开发表的利用带壳和脱壳大麦作为日粮谷物,使用一系列酶制剂的养猪试验报告。尽管很难通过一些因子(如:生长与消化试验、粪与回肠消化率的测定、使用猪的年龄与体重、酶活性的选择、颗粒饲料或粉状饲料,饲料原料变性、使用带壳或脱壳大麦)的潜在影响而得出一个总的看法,但是,趋势还是比较明显的。
仔猪(<25 kg)对这些粘性谷物反应更强烈,研究结果也比较一致,尤其是近些年来研究较多的,饲喂脱壳大麦基础日粮的仔猪的结果一致性更好。Baidoo等(1998a)研究了不同体重的猪对日粮中添加酶制剂的反应,研究表明在超过40-60 kg体重后,猪对日粮添加酶制剂的反应降低,而9-20 kg仔猪对日粮添加酶制剂反应明显(日增重+12%、饲料/增重-3%),体重在20-40 kg的仔猪对日粮添加酶制剂反应更明显(日增重+17%、饲料/增重-10%),甚至在本试验中,体重在40-60 kg的中猪对日粮添加酶制剂反应也比较明显(日增重+4%、饲料/增重-12%),但因为标准差较大,所以未能达到统计上的显著水平。生长育肥猪(体重>25 kg)的研究数据表明,这些猪对日粮中添加酶制剂的反应,有些研究没有反应,有些则有较小的反应(Graham等,1989;Thacker等,1992a,b),还有些研究表明日粮中添加酶制剂,能显著增加日粮的消化率和/或猪的生长率(van Lunen和Schulze,1996a;McCracken等,1996;Ramaswamy等,1996;Baidoo等,1998a,b)。甚至一些表观赏“没有反应”的试验中(Thacker等,1992a),在使用酶制剂后,均能改善粪蛋白的消化率以及对饲料能量产生的很多影响。这表明尤其在生长育肥猪试验中,酶制剂可以潜在性的释放更多的营养成分,但这种释放的养分必须能在改善能量利用率和增加瘦肉。在酶制剂以最大量向配方日粮中添加,其释放的养分可能超过了动物为沉积瘦肉而对营养物质的需要,则胴体参数是对试验结果解释的基本需要。
Baidoo等(1998a)、McCraken等(1996)以及Ham和Froseth(1993)的研究均表明,对大麦基础日粮中添加饲料酶制剂的生产和经济效益反应的关键在于,对谷物质量(能量和氨基酸的可利用率)的评估。因此关键是评估大麦的饲养价值,尤其是对大麦的快速估测。Fairbairn等(1999)描述了很多带壳大麦的对猪营养价值的预测方程,方程以ADF的含量来预测DE含量,R2值为0.85:
DE(kcal/kg,90%干物质基础)=3526 -(92.8×ADF%)。
更复杂的方程包括更多的因子,R2值进一步提高到0.90;如:
DE(kcal/kg,90%干物质基础)=3964 -(81.3×ADF%)-(273.9×酸溶木质素%)-(56.4×总β-葡聚糖含量%)。
近红外分析光谱(NIRS)是快速估测大麦DE(Zijlstra等,1998)和氨基酸含量(Jaikaran等,1998)的最佳技术,会对酶制剂的使用或的最佳成本效率提供足够准确的数据。
但是单独估测DE含量,用于预测对酶制剂的反应需要小心谨慎。在猪日粮中使用小麦的试验(Cadogan等,1999)的经验表明,控制不同饲料样品的采食量与测定的DE关系不大,所以任何最终的筛选方法,均必须考虑到这种反常现象,对关键因子有充分的理解。
小麦、黑小麦和黑麦作为猪日粮中的谷物来源 van Barneveld(1999)综述了小麦对猪的饲养价值方面的文献,发现所有研究的小麦品种的DE含量的变异范围在3.7 MJ/kg(DM)。报道的最低值为13.3 MJ/kg(DM),最高值为17.0 MJ/kg(DM)。与其他谷物一样,小麦也会受到种植方式、品种、生长地区、位置和年份的影响。很多研究表明“次等”和“优等”小麦的DE值差异在0.8-1.4 MJ/kg(DM)之间。
在英国有人研究了不同品种的小麦对饲养价值的影响(Schulze等,1997),试验采用了6个生长在同一地区的小麦品种(以消除生长环境的影响),作为仔猪(初始体重9 kg)日粮中的唯一谷物(表8.2)。在3周的试验里,生长速度和采食量(70-80 g/d)均有不同变化。采食量(体积)和表观DE进食量(DEI;饲料采食量×测定的粪DE)受制粒后配合饲料的持水力(WHC)的影响,方程式如下:DEI(kcal/d)=3325.1 - 663.64×WHC(g/g);(p=0.015,r2=0.81)。这表明日粮纤维(可溶性+不可溶性纤维)含量是主要影响DEI(DE进食量)的因素。这与Kyriazakis和Emmans(1995)的研究一致,后者研究表明,纤维类型的变化可以通过增加配合饲料的WHC,而影响仔猪(12-25 kg)的采食量(图8.4)
表8.2 不同小麦品种的饲养价值(Schulze等,1997)
a:所有的小麦品种生长在同一地区;b:实测的可消化能;WHC:持水力
Cadogan等(1999)以断奶仔猪(7.5-15.6 kg)研究了刚收获和贮藏10个月后的10个澳大利亚小麦品种的饲养价值。研究表明饲喂10个品种的小麦猪的生长性能变异范围为233-447 g/d(平均值388,标准差62.9),显著受到饲料采食量(平均值438 g/d,标准差70.0,变异范围271-514 g/d)的影响。10个小麦品种的表观DM消化率与猪的生长性能无关,这表明用粪消化率预测猪生产性能有一定的局限性,研究没有给出影响采食量的其他因子。经过10个月的贮藏后,尽管采食量和平均日增重平均比10个月前分别提高了11.3%和9.8%,但是小麦品种之间的显著不同情况仍然存在。
关于黑小麦和黑麦对猪饲养价值变异研究的较少。Van Barneveld(1999)引用Charmley和Greenhalgh(1987)的数据,后者的研究表明黑小麦的饲养价值(15.1 MJ/kg DM)低于小麦(16.0 MJ/kg DM),但是这种不同与其研究的3个黑小麦品种之间的差异相比还是较小的。似乎黑小麦和黑麦饲养价值的变异最起码与小麦一致,两种饲料原料均含有较高水平的非淀粉多糖,尤其是可溶性木聚糖(Cyran等,1995)。用黑麦基础日粮来饲喂家禽,因其能增加食糜粘度,影响营养物质的消化和禽的生长,因此对禽类来说是一个挑战。以同样量饲喂猪因为生理原因可能不存在此问题,但是黑麦的饲养价值仍低于小麦,可能是因为其纤维含量过高(Bayalo,1990)。
猪日粮中小麦、黑小麦和黑麦对外源性酶制剂的反应 小麦、黑小麦和黑麦的胚乳和糊粉层中的主要非淀粉多糖(NSPs)是木聚糖,木聚糖能占到总NSPs的50%-60%(Dierick和Decuypere,1994;Eveers等,1999),这与大麦和燕麦(其主要的NSPs是β-葡聚糖)不同。在胚乳的瘦小细胞壁中的木聚糖的水溶性因品种、生长环境等的不同而不同,而糊粉层中肥大的细胞壁的木聚糖主要是不可溶的。因此对此类谷物及其副产品(如小麦麸、次粉等)来说,外源性木聚糖酶的使用比较重要。表8.3总结了过去10年里公开发表的,以小麦、黑小麦和黑麦配制的基础日粮,添加不同酶制剂后对猪生产性能的影响。
与表8.1一样,一般以仔猪(<25 kg)作试验,用生长育肥猪的试验较少。这是基于幼龄动物每天的营养(能量和氨基酸)进食量,是限制其发挥最佳瘦肉生长潜力的关键,因此此期对酶的潜在反应更明显。幼龄猪日粮中添加酶制剂改善了猪的生产性能(生长率、饲料/增重或消化率)的研究文献更多一些,但是这种趋势受到一个事实的影响,这个事实是极少有研究在添加一种适当的酶之前,预先确定小麦/黑小麦/黑麦的饲养价值。在最近的研究中有人尝试了这项研究(Choct等,1999;Partridge等,1999),按“优质”小麦上已经证实的添加量添加木聚糖酶,表明显著改善了饲喂“次等”小麦的猪的生产性能,与猪的年龄无关(表8.4)。
可溶性戊聚糖对木聚糖酶反应的相对重要性,在以黑麦为主要谷物的猪日粮的研究中受到质疑(Thacker等,1991,1992a;Bedford等,1992;Thacke和Bass,1996)。Dusel等(1997b)研究表明,复合谷物(高浸出粘性的小麦、黑麦和大麦)的猪基础日粮对酶制剂的反应已经远远超出了粘度降低带来的益处(表8.5)。正如Choct等(1999)和Partridge等(1999)的研究,使用木聚糖酶或者木聚糖酶与蛋白酶混合使用,表观上限制了猪的采食量,因此生产性能的改善是因为营养物质的可利用率和进食量增加的结果。其他方面较微妙的微生物群落的变化可能也影响动物对酶添加的反应。添加木聚糖酶处理的猪小肠细菌繁殖减少(图8.6),表明一定程度上营养“节俭”可能也有助于改善生产性能。所有的这些研究表明除非同时测定很多参数,否则很难解释动物对外源性酶有或没有反应。
表8.4以已知饲养价值(Partridge等,1999)的小麦配制的猪(28-60 kg)日粮中,添加木聚糖酶,对猪生产性能的影响
a,b:同一列上标字母不相同者表示差异显著(p<0.05);
*:p<0.05;NS:不显著。
表8.5 酶制剂对饲喂高浸出粘度小麦基础日粮的仔猪(10-25kg)生产性能的影响(Dusel等,1997b),粘度值系个人交流数据
a,b:同一列上标字母不相同者表示差异显著(p<0.05)
*:低=1.3 mPa s;高=3.3 mPa s
图8.6 饲喂猪高粘性(添加或不添加木聚糖酶/蛋白酶)和低粘度小麦,对猪胃肠道食糜中微< |

















 京公网安备 11010802025824号
京公网安备 11010802025824号